From 2014.igem.org
(Difference between revisions)
|
|
| Line 484: |
Line 484: |
| | <br><br> | | <br><br> |
| | <h1>A visualization of the star peptide- the key variable is the distance between the active sites of the two key enzymes. We were asked to propose a relationship between this distance and reaction rate.</h1> | | <h1>A visualization of the star peptide- the key variable is the distance between the active sites of the two key enzymes. We were asked to propose a relationship between this distance and reaction rate.</h1> |
| | + | |
| | + | <img src="https://static.igem.org/mediawiki/2014/6/64/Oxford_star_peptide.jpg" style="float:right;position:relative; width:100%;margin-left:0%;margin-right:0%;" /> |
| | </div> | | </div> |
| | </div> | | </div> |
Revision as of 15:27, 19 September 2014
#list li {
list-style-image: url("https://static.igem.org/mediawiki/2014/6/6f/OxigemTick.png"); }
}

Alternatives to micro-compartments
Collaboration with Melbourne iGEM team
The star peptide- an introduction
The degradation pathway of DCM by DcmA produces a number of intermediates. A number of these, such as formaldehyde, are suspected to be toxic to our host bacteria above certain concentrations. In addition to expressing enzymes which break down these intermediates, we are considering two different approaches to further minimize the accumulation of these compounds. Alongside the original idea of using micro-compartments, we have also collaborated with UniMelb iGEM and considered attaching our different enzymes to the arms of a star peptide (link to Melbourne wiki here). In doing so, this increases the likelihood of our intermediates, produced by one enzymatic reaction, interacting with the next enzyme in the reaction pathway thereby limiting their accumulation.
Our reaction

Our reaction
Our reaction pathway is as follows:
Reaction 1: DCM + DcmA --> toxic intermediate (formaldehyde)
Reaction 2: toxic intermediate (formaldehyde) + FdhA --> neutral product (formate)
The Melbourne star peptide length can be varied in order to control the rate of reaction. Melbourne, however, were unsure of the form of the relationship between these length and rate and asked Oxford to develop a model of this. We hypothesized that, broadly speaking, the effect of increasing the length of the tether would result in a decrease in reaction rate due to two factors- the increased time taken to diffuse from one enzyme to the other and the decreased likelihood of colliding with an enzyme as the distance travelled increases.
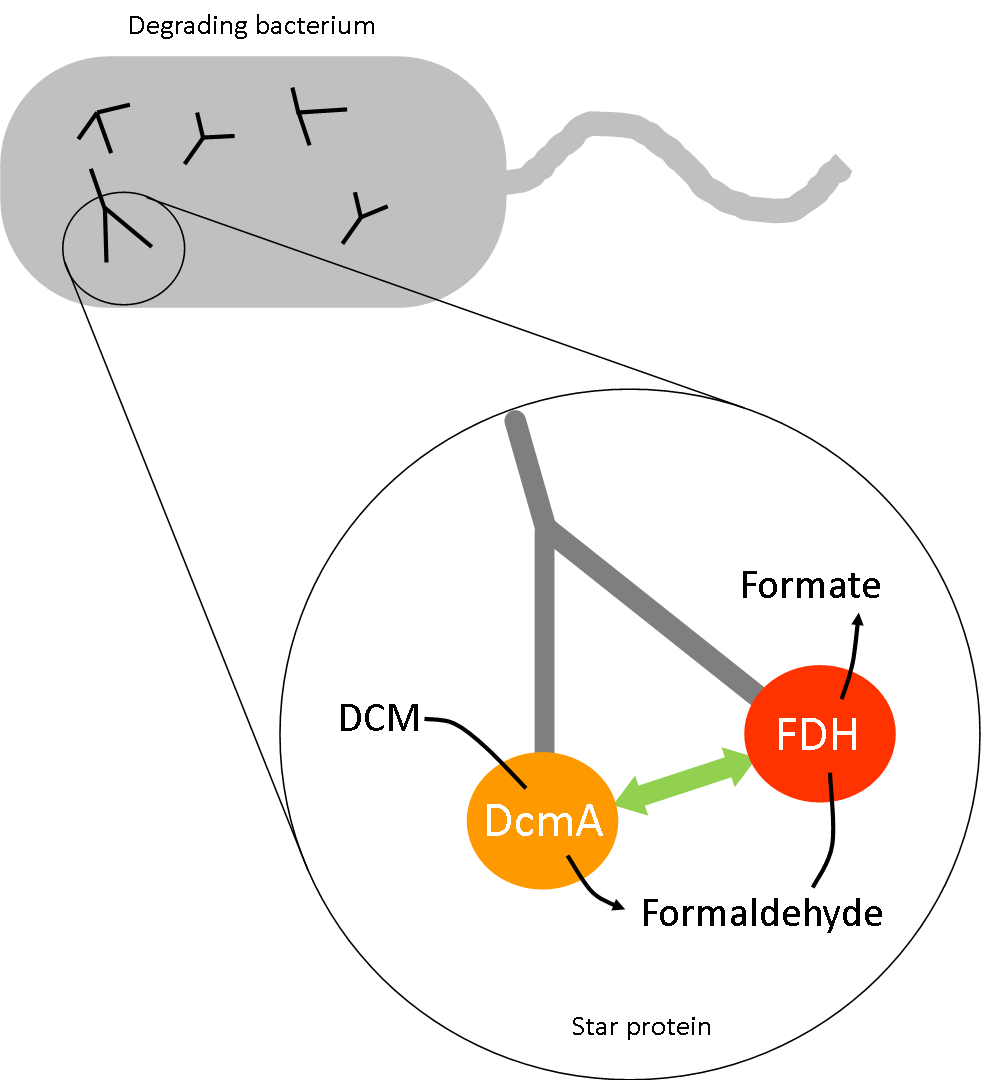
A visualization of the star peptide- the key variable is the distance between the active sites of the two key enzymes. We were asked to propose a relationship between this distance and reaction rate.
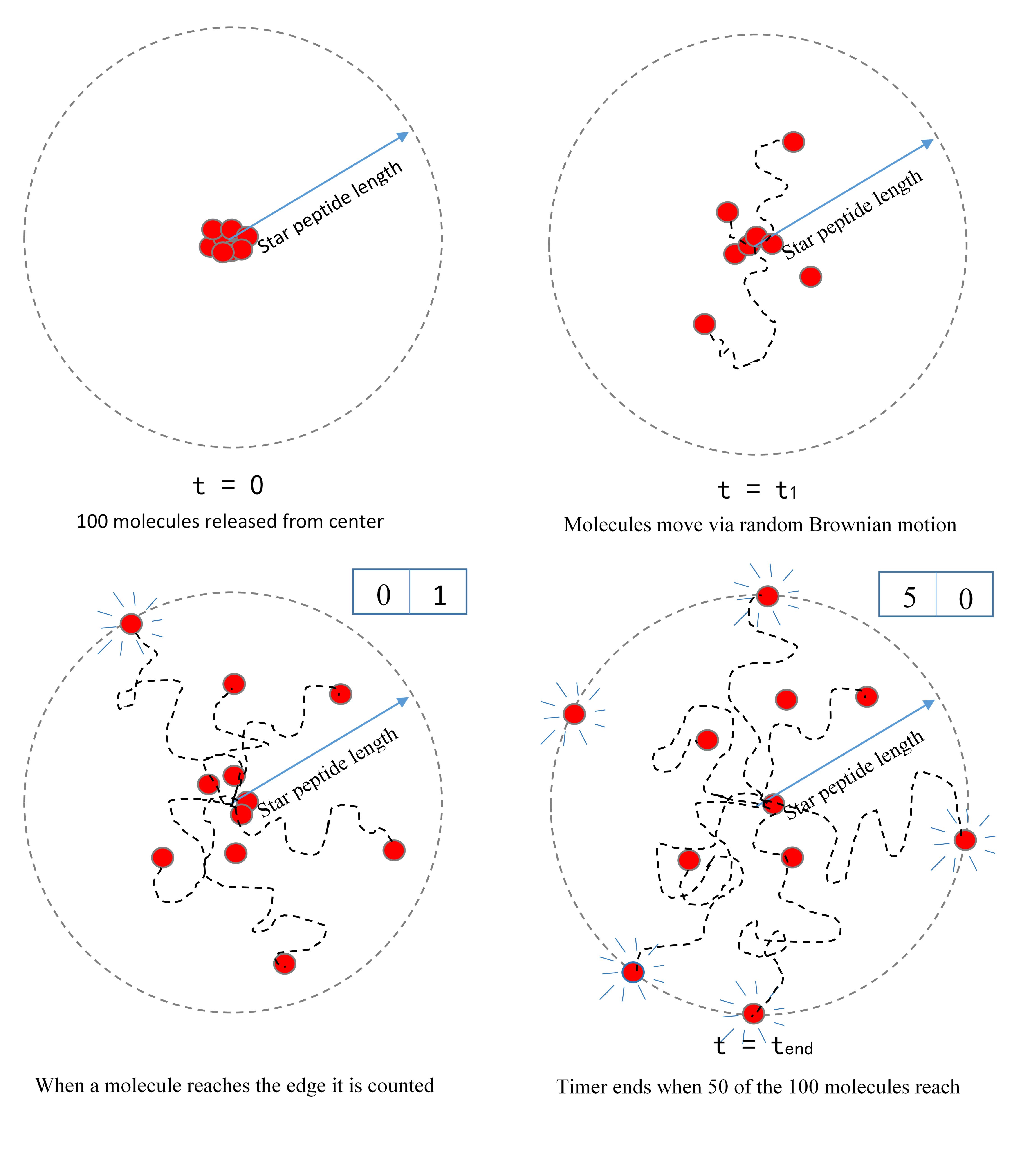
The model
The model
The star peptide- building the model
To isolate the effect of increased diffusion time, our simulation released 100 molecules from the same starting point and tracked their motion through stochastically driven diffusion. It then tracked the time required for 50 of these molecules to reach radii of varying length.
This is again based on another implicit assumption – that a collision between an enzyme and a substrate is a necessary but not sufficient condition for reaction to occur. Therefore, we have assumed that reaction rate is proportional, but not equal to, collision rate. As such, we cannot state the exact reaction rate and have normalized our data so that our highest data point takes a value 1 and subsequent reaction rates are expressed as fractions of this.
The star peptide- model results
The resulting relationship suggested that diffusion rate (defined as 1⁄t_end ) was proportional to 1⁄r^2 . In addition to this, we predict that the relative likelihood of a substrate colliding will an enzyme will decrease as the distance between the enzymes’ active sites increase. This will likely follow a 1⁄r^2 relationship if we consider the fact the likelihood of collision is inversely proportional to the fraction of the surface area of a sphere of the distance r that the enzyme occupies. Thus, we predict that the overall rate-distance relationship will take the form:
reaction rate ∝ 1⁄(peptide length)^4
Thus, the smaller the distance of separation, the higher we expect the rate of reaction to be. However, we must note that this model does not take into consideration stearic hindrances and instabilities that set in when the peptide is made too small. Furthermore, the model is only valid for a minimum radius which is defined as the sum of the two enzyme radii.
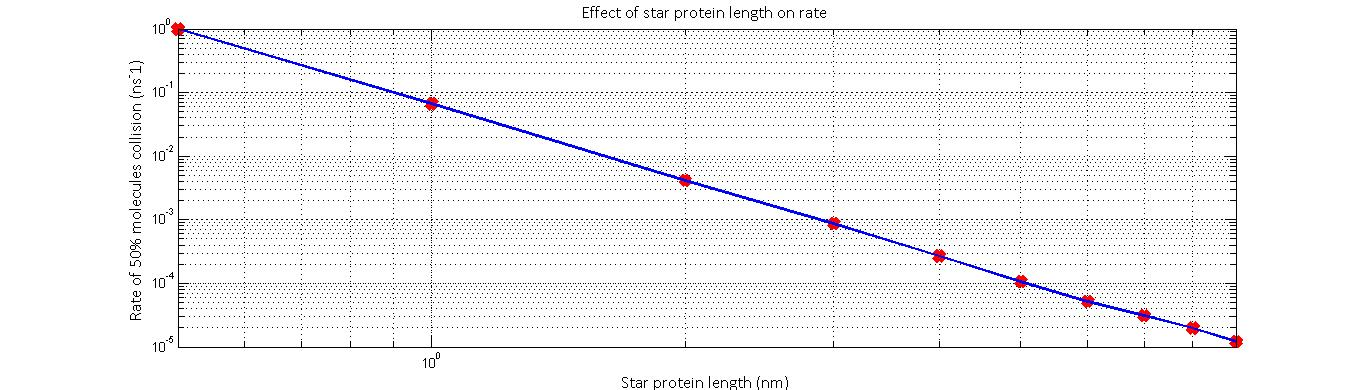
The normalized rate against star protein length results yielded by the model imply that reaction rate is proportional to peptide length^-4.































 "
"















