 "
"
Team:Oxford/what are microcompartments
From 2014.igem.org

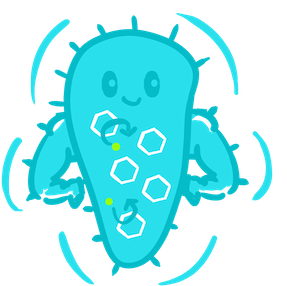


Introduction: what are microcompartments?
In order to produce an efficient bioremediation system, we had to think outside the box and not only improve the expression and function proteins we were working with, but optimise the bacteria themselves for the degradation of toxic compounds. We found that microcompartments, which are explained on this page, are ideally suited for our purposes.
 Microcompartments are proteinaceous capsules that have only recently been recognised to exist in a wide range of bacteria. They contain enzymes required for a particular metabolic process. The carboxysome is a particularly well-studied example of a specialised microcompartment, and is shown here as a schematic diagram (S. Frank et al, 2013).
Microcompartments are proteinaceous capsules that have only recently been recognised to exist in a wide range of bacteria. They contain enzymes required for a particular metabolic process. The carboxysome is a particularly well-studied example of a specialised microcompartment, and is shown here as a schematic diagram (S. Frank et al, 2013).
The Pdu microcompartment that we are using in our project is composed of several proteins encoded in an operon consisting of the genes pduA, -B, -T, -U, -N, -J, -K. These are expressed in various stoichiometries to form different polyhedral shapes with a diameter of 200-250 nm. The faces of the polyhedron are formed by the hexagonal shell proteins PduA, PduB, and PduJ, while PduN is thought to form the vertices (Joshua B. Parsons et al, 2010).
We are expressing the pdu-ABTUNJK codon in E. coli in the pUNI vector, shown below, which we received from the University of Dundee iGEM team.

Additionally, we are expressing pduABUTNJK in P. putida, which to our knowledge has not been done before. For this purpose, we transferred the ABTUNJK sequence into the pBBR1MCS vector, shown below:

Oxford iGEM 2014
The pdu-encoded microcompartments which we are introducing into our bacteria are very similar in shape and structure to carboxysomes, so we used their structure as a starting point for predicting the microcompartment structure. Thus, our initial carboxysome-based structure was a perfect icosahedron with diameter 120nm as established from literature. Following this, we further developed the model by noting that many of the images taken of pdu microcompartments suggest slight and random deviations from an icosahedron.
Taking this into consideration, we have developed a simulation which generates pdu microcompartment structures by starting with a regular icosahedron and randomly distorting the location of each co-ordinate by up to 5% of total edge length.
Given more time and information, this model could be further improved by studying as many microcompartments as possible and refining the size of the deviation from a perfect icosahedron. The magnitude of structural deformation likely follows a normal distribution for which a mean and standard deviation could be established.

Taking this into consideration, we have developed a simulation which generates pdu microcompartment structures by starting with a regular icosahedron and randomly distorting the location of each co-ordinate by up to 5% of total edge length.
Given more time and information, this model could be further improved by studying as many microcompartments as possible and refining the size of the deviation from a perfect icosahedron. The magnitude of structural deformation likely follows a normal distribution for which a mean and standard deviation could be established.
An example microcompartment structure (blue) visualized alongside a perfect icosahedral structure (red).
The number of microcompartments in a given cell can vary broadly, from a few microsomes to numbers that occupy the vast amount of the cytosol. Because of their size and the number of protein components, microcompartments are an extensive strain on the metabolism of cells. We have placed the E. coli ABTUNJK codon under control of the T7 promoter, while that for P. putida is placed under control of the T3 promoter.
The aim of this model was to predict a theoretical maximum number of enzyme molecules that can be packed into a single microcompartment. To get a first estimate, without taking into consideration whether this volume of protein would interrupt the biological processes in the cell, we approached the problem volumetrically.
Due to the complexity of the enzyme movements and their interactions, we simplified their structures by approximating them as ellipsoids with axes lengths calculated through modelling the monomers and predicting the structures of the FdhA tetramer and DcmA hexamer respectively.

Oxford iGEM 2014
Due to the complexity of the enzyme movements and their interactions, we simplified their structures by approximating them as ellipsoids with axes lengths calculated through modelling the monomers and predicting the structures of the FdhA tetramer and DcmA hexamer respectively.
Oxford iGEM 2014
The ellipsoid packing problem
 Once treated as ellipsoids, the problem was then reduced to the classical ‘sand packing’ problem. Because the dimensions of these proteins was substantially smaller than the icosahedron (by approximately a factor of 20 in every dimension), we assumed that the geometry of the container i.e. the microcompartment, was not significant.
Once treated as ellipsoids, the problem was then reduced to the classical ‘sand packing’ problem. Because the dimensions of these proteins was substantially smaller than the icosahedron (by approximately a factor of 20 in every dimension), we assumed that the geometry of the container i.e. the microcompartment, was not significant.
Another assumption made in these calculations was that the enzymes could be treated as homogenous. They are of very similar dimensions, varying by no more than 20-30% on any axis, and also have very similar sphericities- the key variable in determining the packing efficiency of the molecules. Sphericity is defined as:

For ellipsoids, a surface area approximation was used:


After calculating the sphericities of the enzymes, the porosity of the system could then be determined through empirical data from literature. Because the DcmA and FdhA sphericities were very similar (0.953 and 0.981 respectively), we considered the system to be composed of a homogenous spheroid species of porosity 0.973 i.e. the weighted average of the two species.
The image in the top right is a visualization of the ellipsoids packing in the microcompartment
The graph on the right shows the empirical ellipsoid packing efficiencies as defined by sphericity
Results
Because enzyme-enzyme interactions are very difficult to predict, and it is difficult to assign an analogous friction factor to their movements, we took the average of the predicted porosities across a range of different friction factors and alternative methods to give a predicted porosity of 37.3%.Combining this information with the relative expression rates of the two enzymes, we predict that:
 We should note that these numbers are a first approximation and do not take into consideration the strain placed on the cell involved in expressing this amount of protein. This model is based entirely on volumetric and geometric laws rather than biological ones and should be considered a first assumption and theoretical maximum given optimum conditions.
We should note that these numbers are a first approximation and do not take into consideration the strain placed on the cell involved in expressing this amount of protein. This model is based entirely on volumetric and geometric laws rather than biological ones and should be considered a first assumption and theoretical maximum given optimum conditions.
Proteins can be inserted into the microcompartment when they have an N-terminal targeting sequence.
In order to show that this targeting mechanism works for our constructs, we have constructed a plasmid with sfGFP fused to the N-terminal tag. When transforming this vector into E.coli along with the pUNI-ABTUNJK vector, we observed high-density fluorescence spots in those cells that expressed both the microcompartment and microcompartment-tagged sfGFP. This is shown in the image on the left, whereas the image on the right shows a control sample without the pUNI-ABTUNJK vector, thus expressing sfGFP evenly in the cell.


The details for the experimental procedure can be found here.
The details for the experimental procedure can be found here.
Out of the seven subunits, 4 of them (B, U, N and K) are his-tagged, which allowed us to detect expression of these subunits, as shown in the western blot image below:

The Western Blot clearly demonstrates successful expression of subunits B, U, N and K in E.coli, but not in Pseudomonas putida. The negative control was a Pseudomonas putida cell lysate without previous transformation. We used the standard western blot protocol described in the downloadable pdf-document at the top of this page.
The Western Blot clearly demonstrates successful expression of subunits B, U, N and K in E.coli, but not in Pseudomonas putida. The negative control was a Pseudomonas putida cell lysate without previous transformation. We used the standard western blot protocol described in the downloadable pdf-document at the top of this page.










Oxford iGEM 2014






















































Retrieved from "http://2014.igem.org/Team:Oxford/what_are_microcompartments"