 "
"
Team:HokkaidoU Japan/Projects/H Stem
From 2014.igem.org
| (58 intermediate revisions not shown) | |||
| Line 1: | Line 1: | ||
| - | {{Team:HokkaidoU_Japan/ | + | {{Team:HokkaidoU_Japan/LargeDropDown/CSS}} |
| - | + | {{Team:HokkaidoU_Japan/Top_Header/CSS}} | |
| - | {{Team:HokkaidoU_Japan/ | + | |
{{Team:HokkaidoU_Japan/Footer/CSS}} | {{Team:HokkaidoU_Japan/Footer/CSS}} | ||
{{Team:HokkaidoU Japan/Modelling/CSS}} | {{Team:HokkaidoU Japan/Modelling/CSS}} | ||
| - | {{Team:HokkaidoU_Japan/ | + | {{Team:HokkaidoU_Japan/CSS}} |
| - | + | ||
| + | |||
<html> | <html> | ||
| + | <div id="background"> | ||
| + | <link href='http://fonts.googleapis.com/css?family=Special+Elite' rel='stylesheet' type='text/css'> | ||
<link href="//maxcdn.bootstrapcdn.com/font-awesome/4.2.0/css/font-awesome.min.css" rel="stylesheet"> | <link href="//maxcdn.bootstrapcdn.com/font-awesome/4.2.0/css/font-awesome.min.css" rel="stylesheet"> | ||
<!--begin header--> | <!--begin header--> | ||
| - | <div id=" | + | <div id="header-top"> |
| - | <div id="header-top" | + | <img src="https://static.igem.org/mediawiki/2014/b/bb/HokkaidoU_Header_Top.png"> |
| + | </div> | ||
| + | <div id="header-bottom"> | ||
| + | <div id="header-bottom-contents"> | ||
| + | <div class="header-title"> | ||
| + | <div class="header-title-top"> | ||
| + | Projects | ||
| + | </div> | ||
| + | <div class="header-title-bottom"> | ||
| + | H-stem System | ||
| + | </div> | ||
| + | </div> | ||
| + | <div class="header-picture"> | ||
| + | <img src="https://static.igem.org/mediawiki/2014/e/e0/HokkaidoU_Header_cupid.png"> | ||
| + | </div> | ||
| + | </div> | ||
| + | </div> | ||
| + | <!--end header--> | ||
| + | <!--begin menu--> | ||
<div class="box"> | <div class="box"> | ||
<div id="box-wrapper"> | <div id="box-wrapper"> | ||
| + | |||
<ul id="ldd_menu" class="ldd_menu"> | <ul id="ldd_menu" class="ldd_menu"> | ||
<li> | <li> | ||
| - | <span class="menu-list"><a href="https://2014.igem.org/Team:HokkaidoU_Japan"><span style="position:relative; top:10px;">Top</span></a></span><!-- Increases to 510px in width--> | + | <span class="menu-list" ><a href="https://2014.igem.org/Team:HokkaidoU_Japan"><span style="position:relative; top:10px;" >Top</span></a></span><!-- Increases to 510px in width--> |
</li> | </li> | ||
<li class="menu_button"> | <li class="menu_button"> | ||
| Line 21: | Line 42: | ||
<div class="ldd_submenu" style="left:-415px; width:970px;"> | <div class="ldd_submenu" style="left:-415px; width:970px;"> | ||
<ul style="margin-left:20px;"> | <ul style="margin-left:20px;"> | ||
| - | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Overview/ | + | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Overview/Background">Overview</a></li> |
| - | + | ||
<li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Overview/Background">Background</a></li> | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Overview/Background">Background</a></li> | ||
<li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Overview/Achievements">Achievements</a></li> | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Overview/Achievements">Achievements</a></li> | ||
| Line 29: | Line 49: | ||
<li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem">H-stem System</a></li> | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem">H-stem System</a></li> | ||
<li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#Overview">Overview</a></li> | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#Overview">Overview</a></li> | ||
| + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#asRNA_stabilization">Stabilization</a></li> | ||
| + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#Modelling">Modelling</a></li> | ||
| + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#Construct">Construct</a></li> | ||
<li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#How_To_Use">How To Use</a></li> | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/H_Stem#How_To_Use">How To Use</a></li> | ||
| - | |||
</ul> | </ul> | ||
<ul> | <ul> | ||
| - | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034 | + | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034">Anti-sense B0034</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034 | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034">Overview</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034 | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034#Method">Method</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034 | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034#Results">Results</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034 | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/asB0034#Conclusion">Conclusion</a></li> |
| - | + | </ul> | |
<ul> | <ul> | ||
| - | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length | + | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length">Length Variation</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length">Overview</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length#Method">Method</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length#Results">Results</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Projects/Length#Conclusion">Conclusion</a></li> |
</ul> | </ul> | ||
<ul> | <ul> | ||
| Line 55: | Line 77: | ||
</li> | </li> | ||
<li class="menu_button"> | <li class="menu_button"> | ||
| - | <span class="menu-list"> | + | <span class="menu-list"><span style="position:relative; top:10px;">Outreach</span></span> |
<div class="ldd_submenu" style="width:590px; left:-230px;"> | <div class="ldd_submenu" style="width:590px; left:-230px;"> | ||
<ul style="margin-left:20px;"> | <ul style="margin-left:20px;"> | ||
| - | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Univ_festival">Univ. Festival</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Univ_festival#Education">Education</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Univ_festival#Survey">Survey</a></li> |
</ul> | </ul> | ||
<ul> | <ul> | ||
| - | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Survey">High School</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Survey#Education">Education</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Survey#Survey">Survey</a></li> |
</ul> | </ul> | ||
<ul> | <ul> | ||
| - | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_heading"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Discussion">Discussion</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Discussion#Background">Background</a></li> |
| - | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/ | + | <li class="ldd_contents"><a href="https://2014.igem.org/Team:HokkaidoU_Japan/Outreach/Discussion#Evaluation">Evaluation</a></li> |
</ul> | </ul> | ||
</div> | </div> | ||
| Line 88: | Line 110: | ||
</ul> | </ul> | ||
</div> | </div> | ||
| - | |||
</div> | </div> | ||
| - | |||
| - | |||
| - | |||
| - | |||
| - | |||
| + | <!--end box--> | ||
<!-- end menu--> | <!-- end menu--> | ||
<!--begin contents--> | <!--begin contents--> | ||
| Line 105: | Line 122: | ||
<h1 id="Overview">Overview</h1> | <h1 id="Overview">Overview</h1> | ||
<p> | <p> | ||
| - | The weak point of silencing by anti-sense RNA (asRNA) is the instability of RNA. RNA has a shorter half-life than DNA, it is essential to be stabilized | + | The weak point of silencing genes by anti-sense RNA (asRNA) is the instability of RNA. RNA has a shorter half-life than DNA, so it is essential for asRNA to be stabilized to repress mRNA sufficiently. This problem was solved by inserting each-complementary sequeces called stem regions into upstream and downstream of as RNA (N. Nakashima <i>et al.</i>, 2006<sup><a href="#cite-1">[1]</a></sup>). These regions make asRNA form stem-loop structures, which lead to increase the stability of asRNA dramatically. |
</p> | </p> | ||
<p> | <p> | ||
| - | We followed in their footsteps. We designed our | + | We followed in their footsteps. We designed our original stem sequence "H-stem <a href="http://parts.igem.org/Part:BBa_K1524100">(BBa_K1524100)</a>" and produced a BioBricked vector for asRNA expression so that other iGEMers and researchers can easily use stemmed asRNA. By using this vector, you can get an asRNA-expressing construct just by inserting PCRed target gene into the vector. |
</p> | </p> | ||
| Line 116: | Line 133: | ||
<p> | <p> | ||
| - | The | + | The asRNA region is located between two stem regions. (Fig. 1) |
</p> | </p> | ||
| Line 125: | Line 142: | ||
<p> | <p> | ||
| - | Transcripted to RNA, stem | + | Transcripted to RNA, stem sequence binds with each other and make the whole RNA form stem-loop structure. (Fig. 2) |
</p> | </p> | ||
| Line 134: | Line 151: | ||
</div> | </div> | ||
<p> | <p> | ||
| - | This stem-loop structure contributes to RNA | + | This stem-loop structure contributes to stabilize RNA on the following two points. |
</p> | </p> | ||
<p> | <p> | ||
| - | One is the protection from the exonuclease activity of RNase. Since RNase in the cell decomposes single-strand RNA, | + | One is the protection from the exonuclease activity of RNase. Since RNase in the cell decomposes single-strand RNA, stemmed asRNA is hardly affected by such a degradation. |
</p> | </p> | ||
<p> | <p> | ||
| - | The other is an increase of thermodynamical stability by a decrease of Gibbs energy. Stem region produces such a great amount of negative | + | The other is an increase of thermodynamical stability by a decrease of Gibbs energy. Stem region produces such a great amount of negative $\Delta G$ that positive $\Delta G$ produced by the curvature of loop region is overwhelmed. |
</p> | </p> | ||
<div class="clearfix"></div> | <div class="clearfix"></div> | ||
| Line 151: | Line 168: | ||
<p> | <p> | ||
| - | In bacterial cells, the decomposability of asRNA to that of target mRNA affects the amount of translated mRNA as shown below. Here, x-axis | + | In bacterial cells, the decomposability of asRNA to that of target mRNA affects the amount of translated mRNA as shown below. Here, x-axis indicates the transcription level of asRNA to that of mRNA. |
</p> | </p> | ||
| Line 159: | Line 176: | ||
<p> | <p> | ||
| - | From that graph, you can see that the stability of asRNA greatly | + | From that graph, you can see that the stability of asRNA greatly contributes to the repression of mRNA. The detail is discussed below. |
</p> | </p> | ||
| - | <h2 id="Modelling | + | <h2 id="Modelling">Modelling</h2> |
<meta http-equiv="X-UA-Compatible" CONTENT="IE=EmulateIE7"/> | <meta http-equiv="X-UA-Compatible" CONTENT="IE=EmulateIE7"/> | ||
| - | <div | + | <div><!--モデリングで隠せるのはここから--> |
<p> | <p> | ||
| Line 176: | Line 193: | ||
<p> | <p> | ||
| - | Besides, mRNA and asRNA are produced and decomposed at all times. So, the ordinary | + | Besides, mRNA and asRNA are produced and decomposed at all times. So, the ordinary differential equation that represses the whole reaction in the cell is shown below. |
</p> | </p> | ||
| Line 211: | Line 228: | ||
<h1 id="Construct">Construct</h1> | <h1 id="Construct">Construct</h1> | ||
<p> | <p> | ||
| - | + | ||
| + | We constructed a vector "H-stem vector <a href="http://parts.igem.org/Part:BBa_K1524100">(BBa_K1524100)</a>" which includes the H-stem system. | ||
| + | |||
</p> | </p> | ||
| Line 221: | Line 240: | ||
<p> | <p> | ||
| - | + | Stemmed asRNA are translated by a inducible promoter, P<sub>lac</sub> <a href="http://parts.igem.org/Part:BBa_R0010">(BBa_R0010)</a>, and it is activated by IPTG induction. | |
</p> | </p> | ||
<p> | <p> | ||
| - | Since anti-sense region can be digested out by NcoI and XhoI, you can insert any anti-sense sequence as recounted | + | Since anti-sense region can be digested out by NcoI and XhoI, you can insert any anti-sense sequence as recounted above. Additionally, a dummy anti-sense sequence is in advance inserted into this vector. |
| + | </p> | ||
| + | |||
| + | <p> | ||
| + | |||
| + | A point to notice when using our H-stem is that, the stem sequence is difficult to PCR, especially in sequencing PCR. | ||
| + | It is supposed that single-strand DNA of this sequence also forms stem-loop structure and it prevents from the extension of complementary strand, but the detailed mechanism is unknown. | ||
| + | Therefore, if you want to sequence the H-stem, you have to divide it’s sequence into upstream part and downstream part before doing PCR. | ||
</p> | </p> | ||
| Line 231: | Line 257: | ||
<p> | <p> | ||
| - | Now, | + | Now, we explain how to use H-stem vector. |
</p> | </p> | ||
| Line 240: | Line 266: | ||
<br> | <br> | ||
| - | <ol> | + | <ol style="display:block;" type="1"> |
<li>Determine the asRNA binding sequence, which is about 60 bases, from the tail of RBS to the head of CDS. (Please confirm that the sequence does not include NcoI or XhoI site.)</li> | <li>Determine the asRNA binding sequence, which is about 60 bases, from the tail of RBS to the head of CDS. (Please confirm that the sequence does not include NcoI or XhoI site.)</li> | ||
<li>Design the primers which amplify the sequence. Here, add XhoI recognition site (5’-GGG<b>CTCGAG</b>…) to 5’ end of the forward primer and NcoI recognition site (5’-GGG<b>CCATGG</b>…) to that of forward primer.</li> | <li>Design the primers which amplify the sequence. Here, add XhoI recognition site (5’-GGG<b>CTCGAG</b>…) to 5’ end of the forward primer and NcoI recognition site (5’-GGG<b>CCATGG</b>…) to that of forward primer.</li> | ||
| Line 255: | Line 281: | ||
<h3>Reference</h3> | <h3>Reference</h3> | ||
<ol class="citation-list"> | <ol class="citation-list"> | ||
| - | <li id="cite-1">N Nakashima <i>et al.</i> (2006) Paired termini stabilize antisense RNAs and enhance conditional gene silencing in <i>Escherichia coli</i>. Nucleic Acids Res 34: 20 e138</li> | + | <li id="cite-1">N. Nakashima <i>et al.</i> (2006) Paired termini stabilize antisense RNAs and enhance conditional gene silencing in <i>Escherichia coli</i>. Nucleic Acids Res 34: 20 e138</li> |
</ol> | </ol> | ||
| Line 263: | Line 289: | ||
<!--end contents--> | <!--end contents--> | ||
<!--begin footer--> | <!--begin footer--> | ||
| - | <div id="footer-wrapper" style="background-color:# | + | |
| - | <div id="footer- | + | <div id="footer-wrapper" style="background-color: #d92424;"> |
| - | + | <div id="footer-content"> | |
| - | + | <div id="footer-logo"> | |
| - | + | <a href="http://igemhokkaidou.wordpress.com"><img style="height:150px;position:relative;" src="https://static.igem.org/mediawiki/2014/3/39/HokkaidoU_logo_transparent.png"></a> | |
| - | + | ||
| - | + | ||
</div> | </div> | ||
| + | <div id="footer-twitter"> | ||
| + | <a href="http://twitter.com/igem_hokkaidou"><img style="height:150px; position:relative; bottom:0;" src="https://static.igem.org/mediawiki/2014/3/3b/HokkaidoU_Footer_Twitter.png"></a> | ||
| + | </div> | ||
| - | < | + | </div> |
| - | + | </div> | |
</div><!--background--> | </div><!--background--> | ||
| + | <link href='http://fonts.googleapis.com/css?family=Merriweather' rel='stylesheet' type='text/css'> | ||
</html> | </html> | ||
| + | {{Team:HokkaidoU_Japan/JS}} | ||
| + | {{Team:HokkaidoU_Japan/Modelling/JS}} | ||
Latest revision as of 15:20, 9 September 2015


Overview
The weak point of silencing genes by anti-sense RNA (asRNA) is the instability of RNA. RNA has a shorter half-life than DNA, so it is essential for asRNA to be stabilized to repress mRNA sufficiently. This problem was solved by inserting each-complementary sequeces called stem regions into upstream and downstream of as RNA (N. Nakashima et al., 2006[1]). These regions make asRNA form stem-loop structures, which lead to increase the stability of asRNA dramatically.
We followed in their footsteps. We designed our original stem sequence "H-stem (BBa_K1524100)" and produced a BioBricked vector for asRNA expression so that other iGEMers and researchers can easily use stemmed asRNA. By using this vector, you can get an asRNA-expressing construct just by inserting PCRed target gene into the vector.
asRNA stabilization by stem structure
The asRNA region is located between two stem regions. (Fig. 1)

Transcripted to RNA, stem sequence binds with each other and make the whole RNA form stem-loop structure. (Fig. 2)
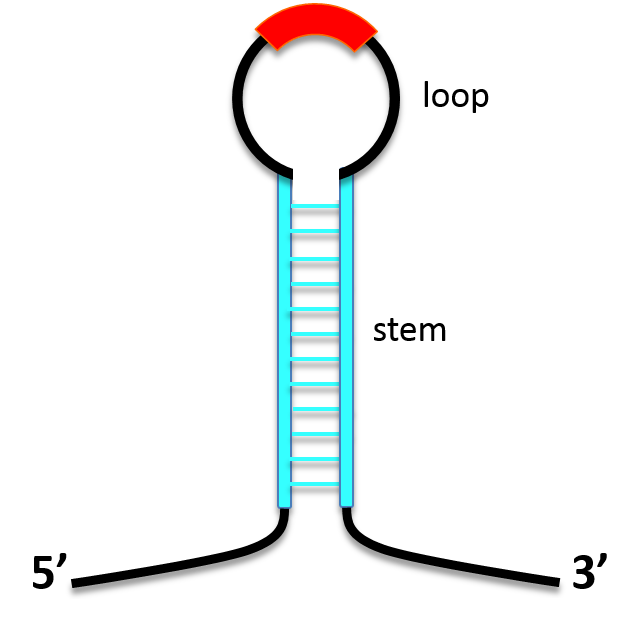
This stem-loop structure contributes to stabilize RNA on the following two points.
One is the protection from the exonuclease activity of RNase. Since RNase in the cell decomposes single-strand RNA, stemmed asRNA is hardly affected by such a degradation.
The other is an increase of thermodynamical stability by a decrease of Gibbs energy. Stem region produces such a great amount of negative $\Delta G$ that positive $\Delta G$ produced by the curvature of loop region is overwhelmed.
In bacterial cells, the decomposability of asRNA to that of target mRNA affects the amount of translated mRNA as shown below. Here, x-axis indicates the transcription level of asRNA to that of mRNA.
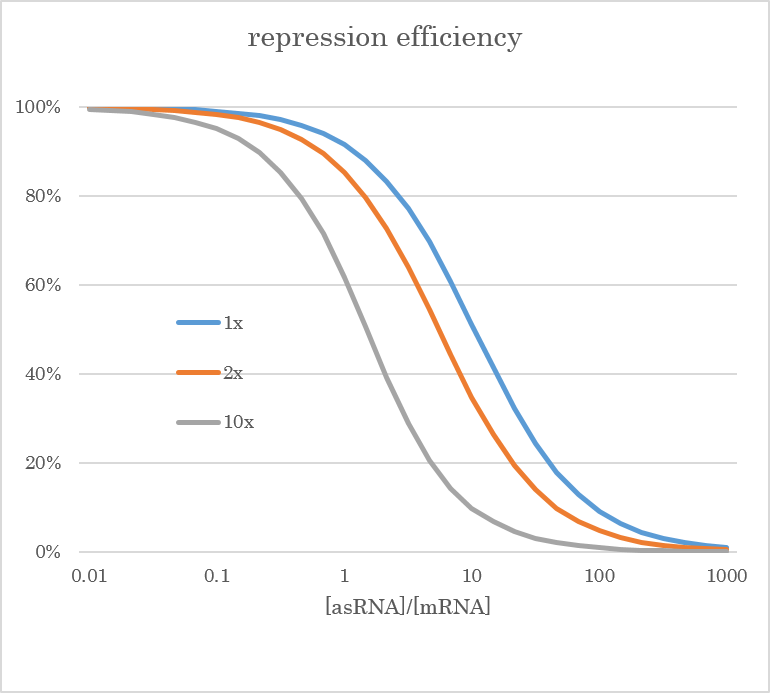
From that graph, you can see that the stability of asRNA greatly contributes to the repression of mRNA. The detail is discussed below.
Modelling
The reaction of asRNA (hereinafter referred to as $X$), mRNA ($Y$) and RNA duplex ($Z$) consisting of the asRNA and the mRNA in a bacterial cell is
\[ X+Y \overset{k_{\rm bind}}{\underset{k_{\rm unbind}}{\rightleftharpoons}} Z \]Besides, mRNA and asRNA are produced and decomposed at all times. So, the ordinary differential equation that represses the whole reaction in the cell is shown below.
\begin{cases} \dot{x}=a-bx-k_{\rm bind}xy+k_{\rm unbind}z & \\ \dot{y}=1-y-k_{\rm bind}xy+k_{\rm unbind}z & \\ \dot{z}=k_{\rm bind}xy-k_{\rm unbind}z-cz & \end{cases}Here, we took $1$ for two constants of the right side first and second terms in the second formula using the flexibilities of $[time]$ and $[concentration]$. Thus, constant $a$ stands for the expression level of asRNA to that of target mRNA and constant $b$ the decomposability of asRNA to that of mRNA.
The unique stable fixed point (in quadrant I) of the ODE is easily found by taking $0$ for each equation. Especially, $y$ value of the fixed point, $y^*$, is
\[ y=\frac{1}{2} \biggl\{ \sqrt{ \bigl( a-1+\frac{b}{\gamma} \bigl)^2 +4 \frac{b}{\gamma}} - \Bigl( a-1+\frac{b}{\gamma} \Bigl) \biggl\} \]Provided that \[ \gamma = \frac{k_{\rm bind}c}{k_{\rm unbind}+c} \]
We find the graph above by regarding this formula as a function of $a$.
Though here we let $\gamma$ value be $1$ arbitarily, this value does not affect qualitatively as the formula shows.
Construct
We constructed a vector "H-stem vector (BBa_K1524100)" which includes the H-stem system.
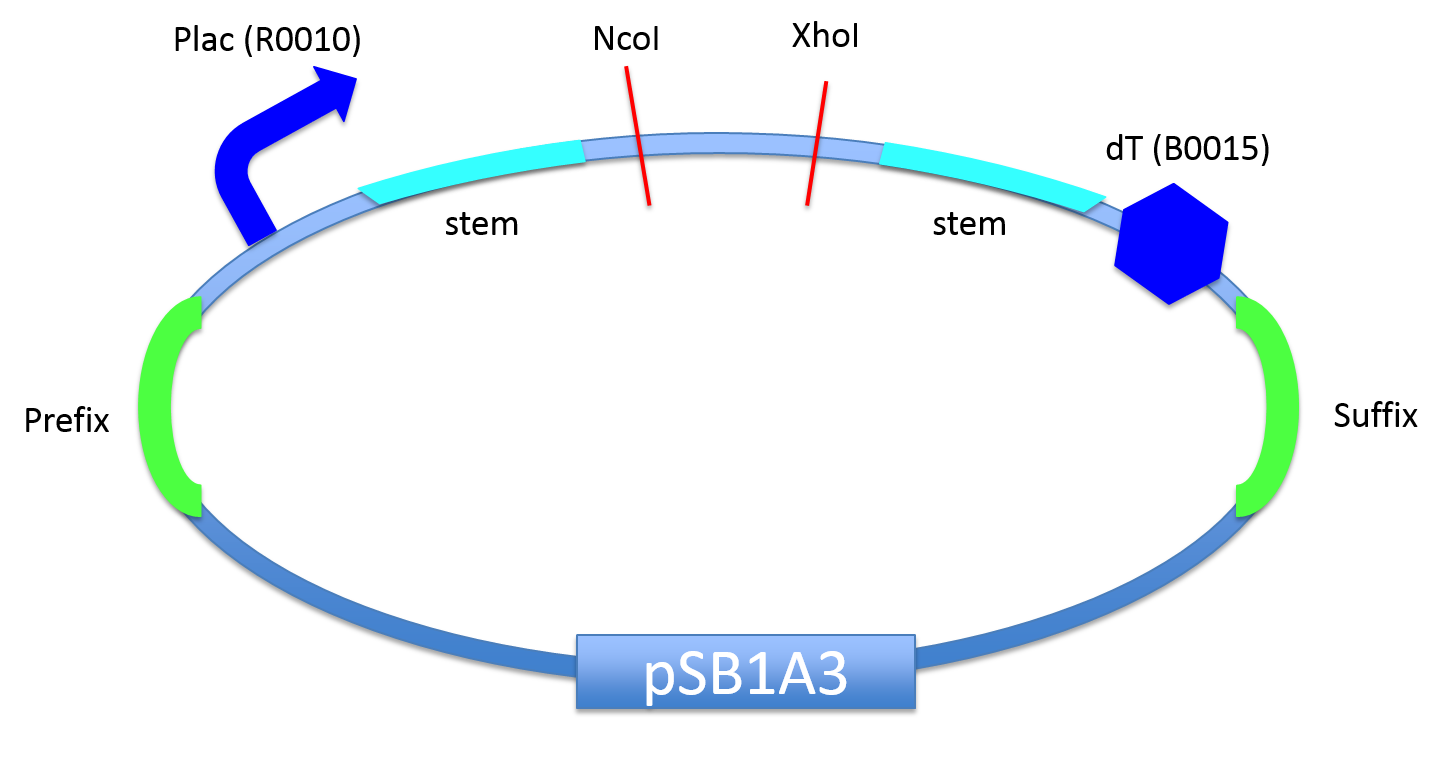
Stemmed asRNA are translated by a inducible promoter, Plac (BBa_R0010), and it is activated by IPTG induction.
Since anti-sense region can be digested out by NcoI and XhoI, you can insert any anti-sense sequence as recounted above. Additionally, a dummy anti-sense sequence is in advance inserted into this vector.
A point to notice when using our H-stem is that, the stem sequence is difficult to PCR, especially in sequencing PCR. It is supposed that single-strand DNA of this sequence also forms stem-loop structure and it prevents from the extension of complementary strand, but the detailed mechanism is unknown. Therefore, if you want to sequence the H-stem, you have to divide it’s sequence into upstream part and downstream part before doing PCR.
How to Use
Now, we explain how to use H-stem vector.
What you need is two restriction enzymes, NcoI and XhoI, and two PCR primers, added to the H-stem vector.
- Determine the asRNA binding sequence, which is about 60 bases, from the tail of RBS to the head of CDS. (Please confirm that the sequence does not include NcoI or XhoI site.)
- Design the primers which amplify the sequence. Here, add XhoI recognition site (5’-GGGCTCGAG…) to 5’ end of the forward primer and NcoI recognition site (5’-GGGCCATGG…) to that of forward primer.
- PCR the sequence by the two primers.
- Digest the PCR product and H-stem vector by NcoI and XhoI, mix them, and ligate together.
We hope that this method will be a help to many iGEMers.
Reference
- N. Nakashima et al. (2006) Paired termini stabilize antisense RNAs and enhance conditional gene silencing in Escherichia coli. Nucleic Acids Res 34: 20 e138
