"
"
Team:ETH Zurich/project/background
From 2014.igem.org

Background
<article class="emergence complex"> </html>
Emergence, Complexity and Simplicity
"The whole is more than the sum of its parts" Aristotle
Emergence describes how novel properties can arise from lower-complexity parts, where these properties are not observed.[12] This phenomenon is wide-spread in nature. One straight-forward example is water: molecules of H2O are here considered as simple subparts. The liquid "water" is a system consisting of several H2O molecules. One well-known property of this system is temperature. Nevertheless, no molecules can be characterized by such a property. Along with other properties as viscosity and pressure, temperature is an emergent property of the system.
In nature, one particular pattern formation retained our attention: some sea snail present a complex pattern on their shells. The "Cloth of Gold" observed on textile cones results from an inner computation of the neural cells of the sea snail. The complex pattern emerges from the assembly of different cells.

Pattern can be characterized by their complexity. Complexity is no synonym for randomness or for complication. Complexity absorbs two a priori opposite concepts: order and randomness. A complex pattern is random and ordered. How is it possible to merge the two concepts together? How can a scientist take it into account? Our Human Practice section is focused on those philosophical and epistemiological issues.
Emergence, Complexity and Simplicity
"The whole is more than the sum of its parts" Aristotle
When one considers an object, it is always made of sub-parts. For example, a human is made of cells. A table is made of molecules. Each sub-part has some properties. Nonetheless, the object has other properties. These global properties can, more or less directly, be linked to the sub-part properties. This is called Emergence. Another way to see it is that an object is more than only the juxtaposition of sub-parts. For instance, interactions between sub-parts have a huge influence on the property of the system.
Water is made of H20 molecules. Water can be hot or cold. The temperature is a property of water. However, a H2O molecule has no temperature. It is not one of its property. Only a set of molecules can have a temperature.
In nature, emergence is a common phenomenon. For instance, those sea snails have a colorful shell. Those colors are not completely ordered. We would detect a repetitive motif on the shells. Those colors are not randomly put on the shells. We can recognize some geometrical forms. This mixture of randomness and order is a characteristic called complexity. As complexity is a buzz word in science, we chose to investigate it more in our Human Practice section and throughout our project.
Pattern Formation
Cellular Automata
Pattern formation was formalized by Neuman in the concept of cellular automata. Following a simple pre-programmed logic rule, the state of a cell is determined by the states of three parent cells (from the previous computation round). Those logic rules are discrete computations: they can be either implementing an AND logic gate or another rule. Wolfram [11] elaborated a whole theory on these cellular automata.
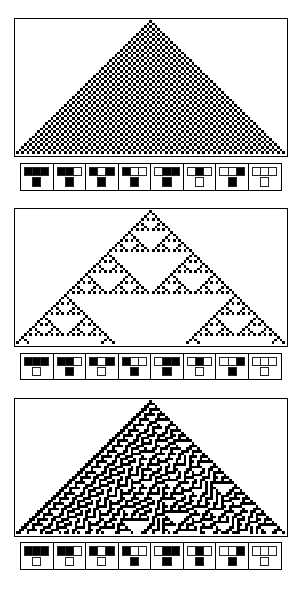
According to cellular automata theory, emergent patterns offer a large panel of properties: striking examples are the rule 30 which gives an apparently random pattern and the rule 110 which has been proven to be Turing complete. With cellular automata, you cannot predict how the final pattern will look like even if you know the rule that governs its property. Thus, the intricated computations of steps poses the problem of complexity.
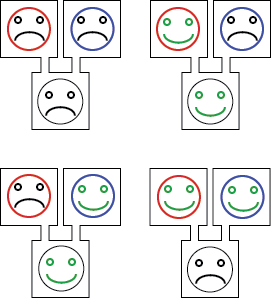
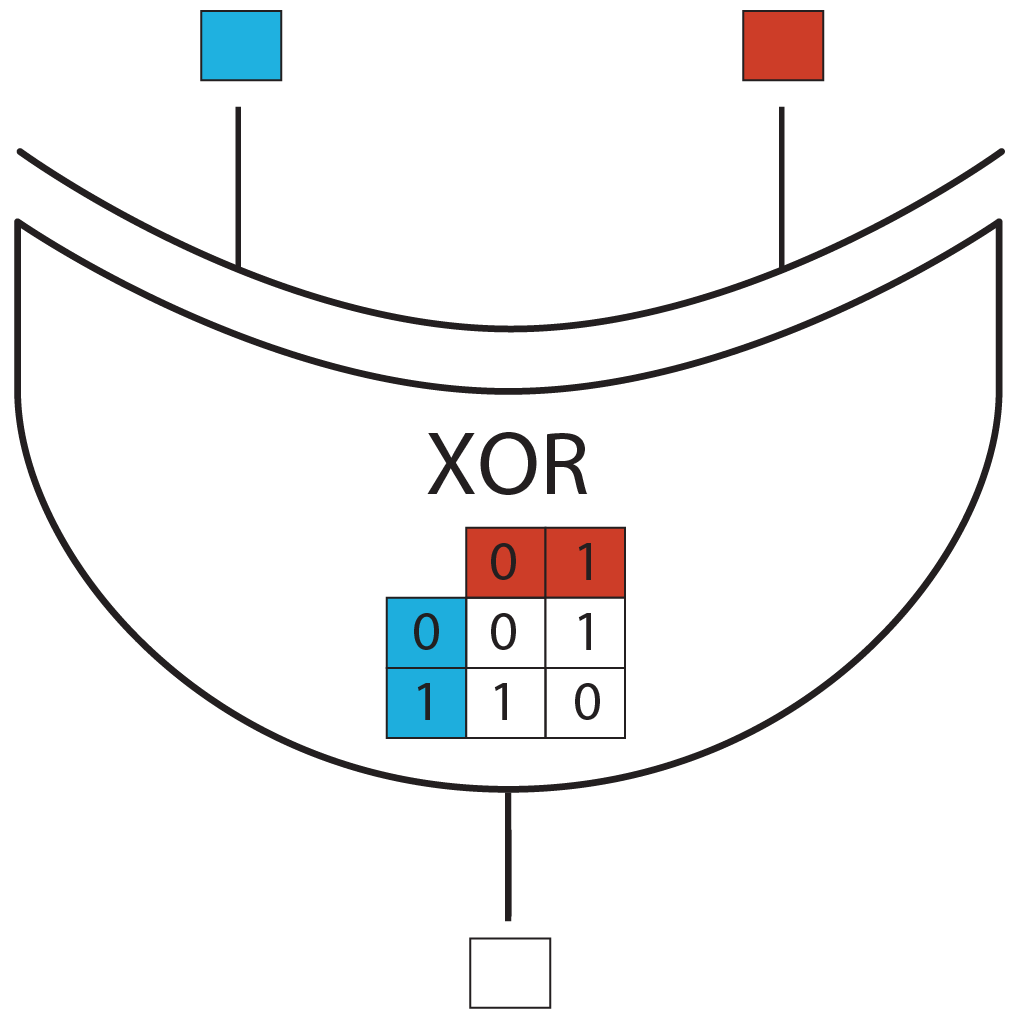
We chose to implement the XOR gate with two inputs, corresponding to rule 6. It can be considered as a first step towards the rule 110, which is an XOR with 3 inputs and is known to be Turing complete.


Biological Tools
Quorum Sensing
Bacteria, usually found in multi-species neighborhoods with billions of cells, where thought to live and divide isolated from its surroundings. However, bacteria do not try to out-compete the neighbors by mindless exponential growth but can exchange information via cell-to-cell communications in order to coordinate gene-expression and as a result the overall behavior of the population. This communicative endeavor is commonly described as quorum sensing and based on the synthesis, secretion and diffusion of small molecules or 'autoinducers'.
One of the best studied quorum sensing models was found in the Gram-negative bacterium V. fischeri in the early 1980s[4]. Depending on the cell density and the corresponding autoinducer concentration, communication between the bacteria induces bioluminescence in the whole V. fischeri population. The small molecule responsible for this behavior was identified as N-(3-oxohexanoyl)-L-homoserine lactone, which is part of the N-acyl homoserine lactone (AHL) family. This AHL is synthesized by an enzyme called LuxI, diffuses through cell membranes and can then reach the cytosol of neighboring cells. There it binds to and activates LuxR, a protein positively regulating gene transcription. Subsequently to the investigation of V. fischeri, several other AHLs (with specific acyl side chains) and corresponding LuxR/LuxI like systems were found in various bacterial species, coordinating diverse responses including plasmid transfer, virulence, and antibiotic resistance[5].
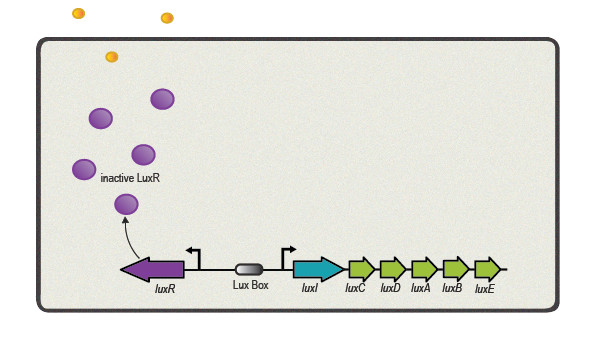
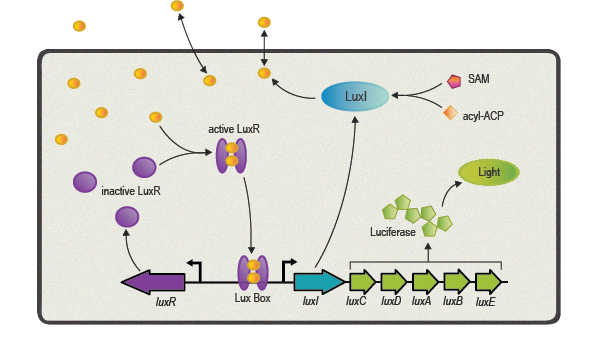
Here we focus in particular on the quorum sensing circuitry of V. fischeri and Pseudomonas aeruginosa, another Gram-negative bacteria. The additional systems are called LasR/LasI and RhlR/RhlI, also employing AHL molecules as autoinducers and are similar to the prototypical LuxR/LuxI system. Even though a key feature of quorum sensing molecules and its receptors is high specificity designed for intraspecies communication, the structural similarity of the different AHL molecules can cause unwanted gene expression. This cross-talk has to be taken into account when re-engineering LuxR/LuxI like systems for synthetic modules and circuits[6].
Integrases
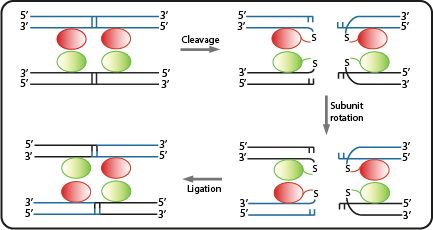
Site specific serine recombinases, or integrases, are enzymes recognizing pairs of short and non-identical DNA sequences. Within those sites they irreversibly catalyze the excision or unidirectional inversion of the DNA bases, depending on the orientation of the recognition sequence (often referred to as attB or attP). Many recombinases originate from phages integrating their genes into bacterial host genomes, hence they often do not require specific co-factors and are fully functional when heterologously expressed in bacteria. As a result, they can be employed as molecular tools in biotechnology[7].
Due to the site-specificity, recombinases like Bxb1 and ΦC31 can be employed orthogonally in synthetic systems. In addition, the combination of multiple recombinases -and their corresponding specific recognition sites- with gene regulatory sequences like promoters and terminators, allowed the engineering of Boolean logic gates[8]. For example, when an asymmetric terminator sequence is placed within a pair of recombination sites, the RNA polymerase can only proceed along this sequence if exactly one recombinase is available. Otherwise the terminator is blocking gene transription by default, or due to the repeated inversion, is again fully functional as soon as both recombinases are present[9]. Such recombinase-based logic gates are at the core of our genetic circuits.